Transcriptomic Analysis of Aedes aegypti Innate Immune System in Response to Ingestion of Chikungunya Virus


Abstract
1. Introduction
2. Results
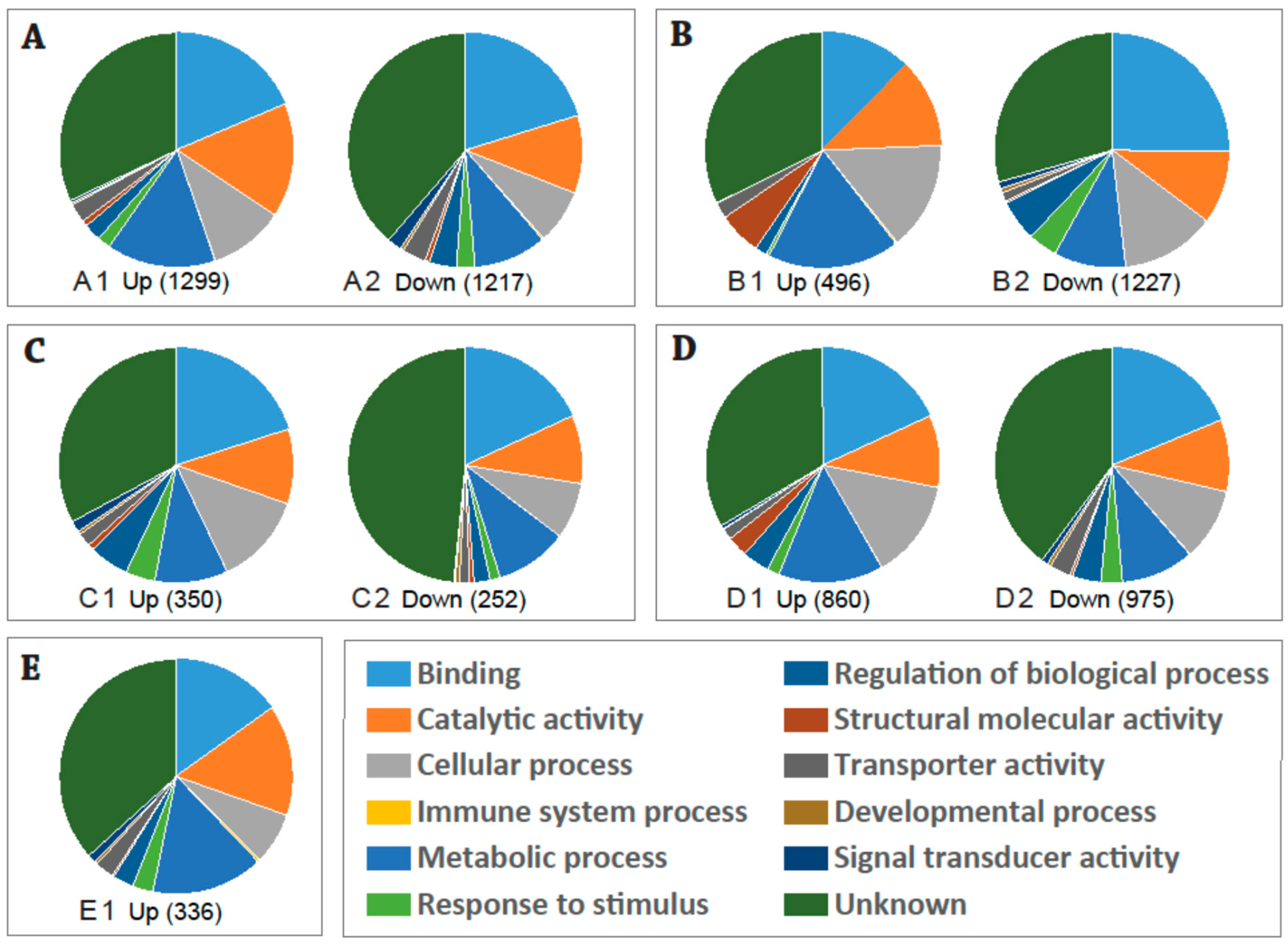
2.1. Global Changes in Transcriptome of the Aedes aegypti Female Adult in Response to CHIKV Infection
2.2. Expression Profiles of DE (Differentially Expressed) Transcripts in Response to Chikungunya Virus (CHIKV) Infection in Aedes aegypti Key West Population/Strain
2.3. Expression Profiles of the DE Transcripts in Response to Blood-Feeding (Control) between the Key West and Orlando Aedes aegypti Strains
2.4. Expression Profiles of the DE Transcripts in Response to CHIKV Infection in the Key West and Orlando Strains of Aedes aegypti
2.5. Expression Profiles of the DE Transcripts in Response to CHIKV Infection in Orlando Aedes aegypti Strain
2.6. Changes of the Immune-Related Genes in the Female Adult of the Key West Strain of Ae. aegypti in Response to CHIKV Infection
2.7. Changes of the Immune-Related Genes in the Female Adult of the Orlando Strain of Ae. aegypti in Response to CHIKV Infection
2.8. Changes of the Immunity-Related Genes of the Female Adults of Ae. aegypti between the Key West and Orlando Strains in Response to CHIKV Infection
2.9. Immunity-Related Genes of the Ae. aegypti Female Adult between the Key West and Orlando Strains in Response to Ingestion of Blood
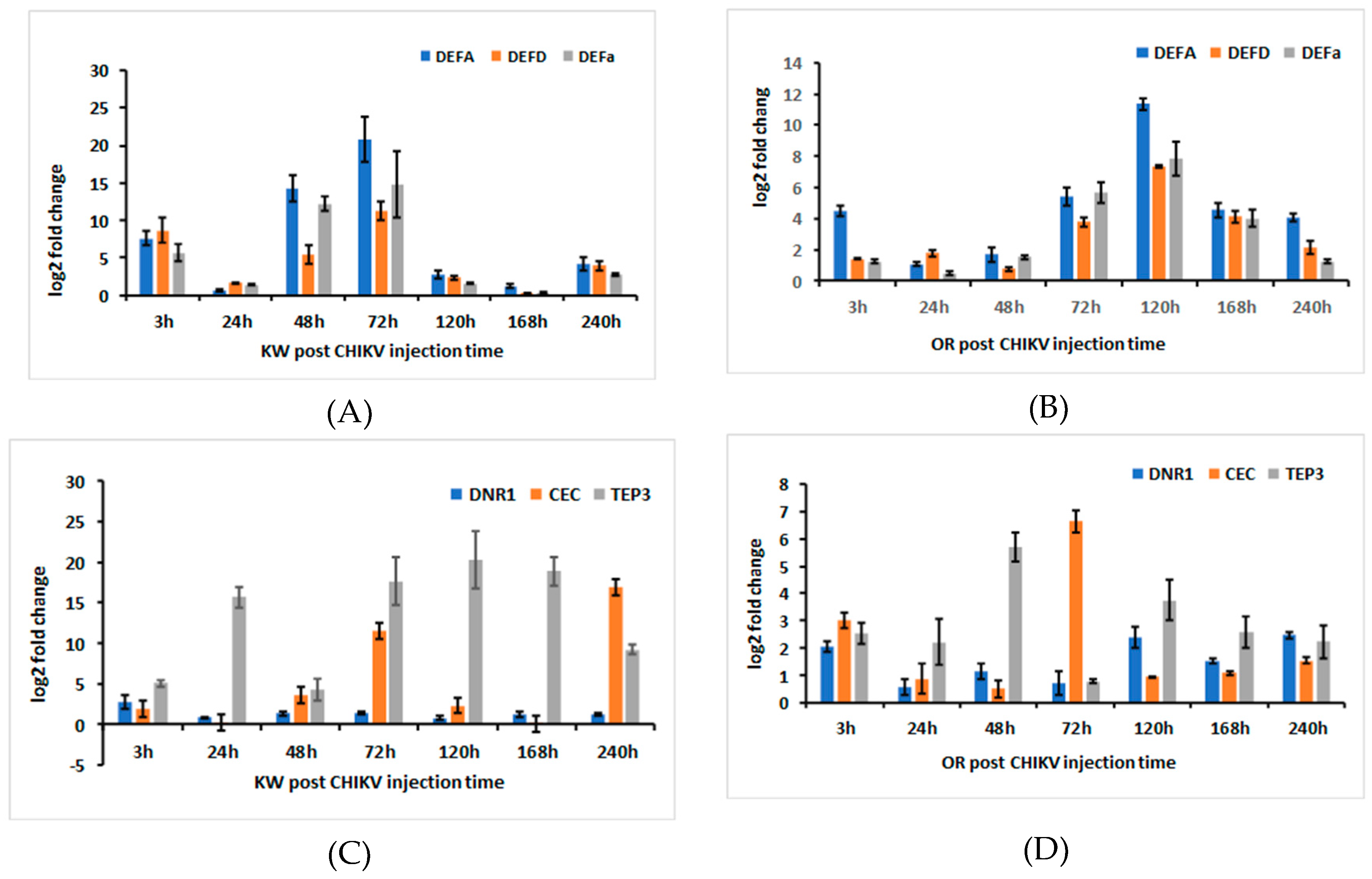
2.10. AaeDEFA, AaeDEFD, AaeDEFa, AaeDNR1, AaeCECH, and AaeTEP3 Transcriptional Induction Following Ingestion of CHIKV-Infected Blood in Ae. aegypti Females
3. Discussion
4. Materials and Methods
4.1. Aedes aegypti
4.2. Chikungunya Virus Infection
4.3. RNA Extraction
4.4. RNA-Sequencing Library Construction and Sequencing
4.5. RNA-sequencing Data Analysis
4.6. Assignments of Gene Ontology (GO) Terms and Pathway Analyses
4.7. cDNA Synthesized and qPCR Amplification
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CHIKV | chikungunya virus |
| CECH | cecropin anti-microbial peptide |
| DEF | defensin |
| TEP | thioester-containing proteins |
References
- Simo Tchetgna, H.; Sem Ouilibona, R.; Nkili-Meyong, A.A.; Caron, M.; Labouba, I.; Selekon, B.; Njouom, R.; Leroy, E.M.; Nakoune, E.; Berthet, N. Viral Exploration of Negative Acute Febrile Cases Observed during Chikungunya Outbreaks in Gabon. Intervirology 2018, 61, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Valadere, A.M. Transcontinental movement of Asian genotype chikungunya virus. Emerg. Infect. Dis. 2014, 20, 1400–1402. [Google Scholar] [CrossRef] [PubMed]
- Leparc-Goffart, I.; Nougairede, A.; Cassadou, S.; Prat, C.; de Lamballerie, X. Chikungunya in the Americas. Lancet 2014, 383, 514. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Chen, R.; Weaver, S.C. Interspecies transmission and chikungunya virus emergence. Curr. Opin. Virol. 2016, 16, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Sudeep, A.B.; Shil, P. (Bigot) mosquito: An emerging threat to public health. J. Vector Borne Dis. 2017, 54, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Ciocchetta, S.; Prow, N.A.; Darbro, J.M.; Frentiu, F.D.; Savino, S.; Montarsi, F.; Capelli, G.; Aaskov, J.G.; Devine, G.J. The new European invader Aedes (Finlaya) koreicus: A potential vector of chikungunya virus. Pathog. Glob. Health 2018, 112, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Powers, A.M. How Chikungunya Virus Virology Affects Its Epidemiology and Transmission: Implications for Influencing Public Health. J. Infect. Dis. 2016, 214, S449–S452. [Google Scholar] [CrossRef] [PubMed]
- Caglioti, C.; Lalle, E.; Castilletti, C.; Carletti, F.; Capobianchi, M.R.; Bordi, L. Chikungunya virus infection: An overview. New Microbiol. 2013, 36, 211–227. [Google Scholar]
- Amdekar, S.; Parashar, D.; Alagarasu, K. Chikungunya Virus-Induced Arthritis: Role of Host and Viral Factors in the Pathogenesis. Viral Immunol. 2017, 30, 691–702. [Google Scholar] [CrossRef]
- Gasque, P.; Bandjee, M.C.; Reyes, M.M.; Viasus, D. Chikungunya Pathogenesis: From the Clinics to the Bench. J. Infect. Dis. 2016, 214, S446–S448. [Google Scholar] [CrossRef]
- Hua, C.; Combe, B. Chikungunya Virus-Associated Disease. Curr. Rheumatol. Rep. 2017, 19, 69. [Google Scholar] [CrossRef] [PubMed]
- Ausubel, F.M. Are innate immune signaling pathways in plants and animals conserved? Nat. Immunol. 2005, 6, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Flajnik, M.F. A cold-blooded view of adaptive immunity. Nat. Rev. Immunol. 2018, 18, 438–453. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Srivastava, P.; Sirisena, P.; Dubey, S.K.; Kumar, R.; Shrinet, J.; Sunil, S. Mosquito Innate Immunity. Insects 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Houk, E.J.; Hardy, J.L.; Chiles, R.E. Permeability of the midgut basal lamina in the mosquito, Culex tarsalis Coquillett (Insecta, Diptera). Acta Trop. 1981, 38, 163–171. [Google Scholar] [PubMed]
- Passarelli, A.L. Barriers to success: How baculoviruses establish efficient systemic infections. Virology 2011, 411, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Behura, S.K. Franz AWE: The midgut transcriptome of Aedes aegypti fed with saline or protein meals containing chikungunya virus reveals genes potentially involved in viral midgut escape. BMC Genomics 2017, 18, 382. [Google Scholar] [CrossRef] [PubMed]
- Kantor, A.M.; Dong, S.; Held, N.L.; Ishimwe, E.; Passarelli, A.L.; Clem, R.J.; Franz, A.W. Identification and initial characterization of matrix metalloproteinases in the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2017, 26, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Bartholomay, L.C.; Cho, W.L.; Rocheleau, T.A.; Boyle, J.P.; Beck, E.T.; Fuchs, J.F.; Liss, P.; Rusch, M.; Butler, K.M.; Wu, R.C.; et al. Description of the transcriptomes of immune response-activated hemocytes from the mosquito vectors Aedes aegypti and Armigeres subalbatus. Infect. Immun. 2004, 72, 4114–4126. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, G.; Richman, A.; Müller, H.M.; Kafatos, F.C. Molecular immune responses of the mosquito Anopheles gambiae to bacteria and malaria parasites. Proc. Natl. Acad. Sci. USA 1997, 94, 11508–11513. [Google Scholar] [CrossRef] [PubMed]
- Bartholomay, L.C.; Fuchs, J.F.; Cheng, L.L.; Beck, E.T.; Vizioli, J.; Lowenberger, C.; Christensen, B.M. Reassessing the role of defensin in the innate immune response of the mosquito, Aedes aegypti. Insect Mol. Biol. 2004, 13, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Bartholomay, L.C.; Michel, K. Mosquito Immunobiology: The Intersection of Vector Health and Vector Competence. Annu. Rev. Entomol. 2018, 63, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, G.; Seeley, D.; Wolf, A.; Kafatos, F.C. Malaria infection of the mosquito Anopheles gambiae activates immune-responsive genes during critical transition stages of the parasite life cycle. EMBO J. 1998, 17, 6115–6123. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, G.; Casavant, T.L.; Chang, S.; Scheetz, T.; Roberts, C.; Donohue, M.; Schultz, J.; Benes, V.; Bork, P.; Ansorge, W.; et al. Anopheles gambiae pilot gene discovery project: Identification of mosquito innate immunity genes from expressed sequence tags generated from immune-competent cell lines. Proc. Natl. Acad. Sci. USA 2000, 97, 6619–6624. [Google Scholar] [CrossRef] [PubMed]
- Osta, M.A.; Christophides, G.K.; Vlachou, D.; Kafatos, F.C. Innate immunity in the malaria vector Anopheles gambiae: Comparative and functional genomics. J. Exp. Biol. 2004, 207, 2551–2563. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Jupatanakul, N.; Ramirez, J.L.; Kang, S.; Romero-Vivas, C.M.; Mohammed, H.; Dimopoulos, G. Transcriptomic profiling of diverse Aedes aegypti strains reveals increased basal-level immune activation in dengue virus-refractory populations and identifies novel virus-vector molecular interactions. PLoS Negl. Trop. Dis. 2013, 7, e2295. [Google Scholar] [CrossRef] [PubMed]
- Etebari, K.; Hegde, S.; Saldaña, M.A.; Widen, S.G.; Wood, T.G.; Asgari, S.; Hughes, G.L. Global Transcriptome Analysis of Aedes aegypti Mosquitoes in Response to Zika Virus Infection. mSphere 2017, 2, e00456-17. [Google Scholar] [CrossRef] [PubMed]
- Shrinet, J.; Srivastava, P.; Sunil, S. Transcriptome analysis of Aedes aegypti in response to mono-infections and co-infections of dengue virus-2 and chikungunya virus. Biochem. Biophys. Res. Commun. 2017, 492, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Alto, B.W.; Shin, D.; Yu, F. The effect of permethrin resistance on Aedes aegypti transcriptome following ingestion of Zika virus infected blood. Viruses 2018, 10, 470. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Civana, A.; Acevedo, C.; Smartt, C.T. Transcriptomics of differential vector competence: West Nile virus infection in two populations of Culex pipiens quinquefasciatus linked to ovary development. BMC Genomics 2014, 15, 513. [Google Scholar] [CrossRef]
- Shrinet, J.; Jain, S.; Jain, J.; Bhatnagar, R.K.; Sunil, S. Next generation sequencing reveals regulation of distinct Aedes microRNAs during chikungunya virus development. PLoS Negl. Trop. Dis. 2014, 8, e2616. [Google Scholar] [CrossRef] [PubMed]
- Bonizzoni, M.; Dunn, W.A.; Campbell, C.L.; Olson, K.E.; Marinotti, O.; James, A.A. Complex modulation of the Aedes aegypti transcriptome in response to dengue virus infection. PLoS ONE 2012, 7, e50512. [Google Scholar] [CrossRef] [PubMed]
- Colpitts, T.M.; Cox, J.; Vanlandingham, D.L.; Feitosa, F.M.; Cheng, G.; Kurscheid, S.; Wang, P.; Krishnan, M.N.; Higgs, S.; Fikrig, E. Alterations in the Aedes aegypti transcriptome during infection with West Nile, dengue and yellow fever viruses. PLoS Pathog. 2011, 7, e1002189. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Smagghe, G.; Meeus, I. The role of a single gene encoding the Single von Willebrand factor C-domain protein (SVC) in bumblebee immunity extends beyond antiviral defense. Insect Biochem. Mol. Biol. 2017, 91, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Alto, B.W.; Smartt, C.T.; Shin, D. Transcription Profiling for Defensins of Aedes aegypti (Diptera: Culicidae) During Development and in Response to Infection with Chikungunya and Zika Viruses. J. Med. Entomol. 2017, 55, 78–89. [Google Scholar]
- Antonova, Y.; Alvarez, K.S.; Kim, Y.J.; Kokoza, V.; Raikhel, A.S. The role of NF-kappaB factor REL2 in the Aedes aegypti immune response. Insect Biochem. Mol. Biol. 2009, 39, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Kokoza, V.; Ahmed, A.; Woon Shin, S.; Okafor, N.; Zou, Z.; Raikhel, A.S. Blocking of Plasmodium transmission by cooperative action of Cecropin A and Defensin A in transgenic Aedes aegypti mosquitoes. Proc. Natl. Acad. Sci. USA 2010, 107, 8111–8116. [Google Scholar] [CrossRef] [PubMed]
- Angleró-Rodríguez, Y.I.; MacLeod, H.J.; Kang, S.; Carlson, J.S.; Jupatanakul, N.; Dimopoulos, G. Molecular Responses to Zika Virus: Modulation of Infection by the Toll and Jak/Stat Immune Pathways and Virus Host Factors. Front. Microbiol. 2017, 8, 2050. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Alto, B.W.; Shin, D. Transcriptional profile of Aedes aegypti Leucine-Rich Repeat Proteins in response to Zika and Chikungunya viruses. Int. J. Mol. Sci. 2019, 20, 615. [Google Scholar] [CrossRef] [PubMed]
- Aloor, J.J.; Azzam, K.M.; Guardiola, J.J.; Gowdy, K.M.; Madenspacher, J.H.; Gabor, K.A.; Mueller, G.A.; Lin, W.C.; Lowe, J.M.; Gruzdev, A.; et al. Leucine-Rich Repeats and Calponin Homology containing 4 regulates the innate immune response. J. Biol. Chem. 2019, 294, 1997–2008. [Google Scholar] [CrossRef]
- Reyes Ruiz, V.M.; Sousa, G.L.; Sneed, S.D.; Farrant, K.V.; Christophides, G.K.; Povelones, M. Stimulation of a protease targeting the LRIM1/APL1C complex reveals specificity in complement-like pathway activation in Anopheles gambiae. PLoS ONE 2019, 14, e0214753. [Google Scholar] [CrossRef] [PubMed]
- Souza-Neto, J.A.; Sim, S.; Dimopoulos, G. An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc. Natl. Acad. Sci. USA 2009, 106, 17841–17846. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Hillyer, J.F. Complement-like proteins TEP1, TEP3 and TEP4 are positive regulators of periostial hemocyte aggregation in the mosquito Anopheles gambiae. Insect Biochem. Mol. Biol. 2019, 107, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Soares, T.S.; Rodriguez Gonzalez, B.L.; Torquato, R.J.S.; Lemos, F.J.A.; Costa-da-Silva, A.L.; Capurro Guimarães, M.L.; Tanaka, A.S. Functional characterization of a serine protease inhibitor modulated in the infection of the Aedes aegypti with dengue virus. Biochimie 2018, 144, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Gaburro, J.; Paradkar, P.N.; Klein, M.; Bhatti, A.; Nahavandi, S.; Duchemin, J.B. Dengue virus infection changes Aedes aegypti oviposition olfactory preferences. Sci. Rep. 2018, 8, 13179. [Google Scholar] [CrossRef] [PubMed]
- Gaburro, J.; Bhatti, A.; Sundaramoorthy, V.; Dearnley, M.; Green, D.; Nahavandi, S.; Paradkar, P.N.; Duchemin, J.B. Zika virus-induced hyper excitation precedes death of mouse primary neuron. Virol. J. 2018, 15, 79. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Ramirez, J.L.; Dimopoulos, G. Dengue virus infection of the Aedes aegypti salivary gland and chemosensory apparatus induces genes that modulate infection and blood-feeding behavior. PLoS Pathog. 2012, 8, e1002631. [Google Scholar] [CrossRef]
- Bonizzoni, M.; Dunn, W.A.; Campbell, C.L.; Olson, K.E.; Marinotti, O.; James, A.A. Strain Variation in the Transcriptome of the Dengue Fever Vector, Aedes aegypti. G3 Genes Genomes Genet. (Bethesda) 2012, 2, 103–114. [Google Scholar] [CrossRef]
- Parola, P.; de Lamballerie, X.; Jourdan, J.; Rovery, C.; Vaillant, V.; Minodier, P.; Brouqui, P.; Flahault, A.; Raoult, D.; Charrel, R.N. Novel chikungunya virus variant in travelers returning from Indian Ocean islands. Emerg. Infect. Dis. 2006, 12, 1493–1499. [Google Scholar] [CrossRef]
- Kim, C.H.; Muturi, E.J. Effect of larval density and Sindbis virus infection on immune responses in Aedes aegypti. J. Insect Physiol. 2013, 59, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Muturi, E.J.; Blackshear, M.; Montgomery, A. Temperature and density-dependent effects of larval environment on Aedes aegypti competence for an alphavirus. J. Vector Ecol. 2012, 37, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.Q.; Yu, F. DEB: A web interface for RNA-seq digital gene expression analysis. Bioinformation 2011, 7, 44–45. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Alto, B.W.; Duguma, D. Transcriptional Profile for Detoxification Enzymes AeaGGT1 and AaeGGT2 from Aedes aegypti (Diptera: Culicidae) in Response to Larvicides. J. Med. Entomol. 2017, 54, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Pridgeon, J.W.; Becnel, J.J.; Clark, G.G.; Linthicum, K.J. Cytochrome c gene and protein expression: Developmental regulation, environmental response, and pesticide sensitivity in Aedes aegypti. J. Med. Entomol. 2008, 45, 401–408. [Google Scholar] [CrossRef]
- Portereiko, M.F.; Sandaklie-Nikolova, L.; Lloyd, A.; Dever, C.A.; Otsuga, D.; Drews, G.N. NUCLEAR FUSION DEFECTIVE1 encodes the Arabidopsis RPL21M protein and is required for karyogamy during female gametophyte development and fertilization. Plant Physiol. 2006, 141, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Portereiko, M.F.; Lloyd, A.; Steffen, J.G.; Punwani, J.A.; Otsuga, D.; Drews, G.N. AGL80 is required for central cell and endosperm development in Arabidopsis. Plant Cell 2006, 18, 1862–1872. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Strains | Initial Dose in Bloodmeal | Freshly Fed | 1 Day Post Infection | 2 Days Post Infection | 5 Days Post Infection | 7 Days Post Infection | 10 Days Post Infection |
|---|---|---|---|---|---|---|---|
| Key West | 8.0 ± 0.09 | 4.89 ± 0.39 | 3.79 ± 0.23 a | 4.47 ± 0.32 ab | 5.87 ± 0.39 bc | 5.83 ± 0.39 bc | 4.96 ± 0.39 ab |
| Orlando | 8.3 ± 0.08 | 5.68 ± 0.39 | 3.09 ± 0.23 A | 1.70 ± 0.30 B | 4.78 ± 0.39 C | 4.02 ± 0.32 AC | 4.24 ± 0.39 AC |
| Transcript ID | Log2 FC | p-adj | Gene Description |
|---|---|---|---|
| AAEL004199-RA | −1.9388 | 3.4 × 10−4 | allergen |
| AAEL003057-RB | 2.8963 | 4.2 × 10−6 | allergen |
| AAEL014658-RA | 2.2755 | 4.4 × 10−4 | caspase-1 protein |
| AAEL000621-RA | 2.2216 | 3.4 × 10−4 | cecropin anti-microbial peptide |
| AAEL005374-RA | −3.2830 | 1.1 × 10−27 | Class B Scavenger Receptor (CD36 domain) |
| AAEL005979-RA | −2.0366 | 1.9 × 10−4 | Class B Scavenger Receptor (CD36 domain) |
| AAEL005981-RA | −1.8895 | 1.5 × 10−5 | Class B Scavenger Receptor (CD36 domain) |
| AAEL000234-RA | 1.8135 | 1.6 × 10−4 | Class B Scavenger Receptor (CD36 domain) |
| AAEL009432-RA | 4.0915 | 3.5 × 10−12 | Class B Scavenger Receptor (CD36 domain) |
| AAEL001098-RA | −2.2546 | 2.9 × 10−7 | Clip-Domain Serine Protease |
| AAEL014137-RA | −2.3522 | 3.2 × 10−7 | Clip-Domain Serine Protease family B |
| AAEL008668-RA | 1.8083 | 1.8 × 10−3 | Clip-Domain Serine Protease family B |
| AAEL000760-RA | 2.2343 | 4.1 × 10−5 | Clip-Domain Serine Protease family B |
| AAEL005648-RA | −1.9389 | 2.0 × 10−3 | Clip-Domain Serine Protease family B |
| AAEL005792-RA | −2.0842 | 5.1 × 10−4 | Clip-Domain Serine Protease family E |
| AAEL001077-RA | −1.9692 | 6.8 × 10−13 | Clip-Domain Serine Protease family B |
| AAEL008299-RA | −2.4009 | 7.4 × 10−5 | C-Type Lectin (CTL) |
| AAEL004679-RA | −2.0008 | 5.5 × 10−3 | C-Type Lectin (CTL) |
| AAEL000533-RA | 2.0222 | 1.9 × 10−4 | C-Type Lectin (CTL) |
| AAEL000556-RA | 3.3807 | 2.4 × 10−28 | C-Type Lectin (CTL) |
| AAEL011078-RA | −2.5784 | 1.9 × 10−3 | C-Type Lectin (CTL)—galactose binding |
| AAEL011455-RA | 2.3166 | 2.9 × 10−4 | C-Type Lectin (CTL)—mannose binding |
| AAEL009384-RA | −1.9456 | 3.7 × 10−7 | fibrinogen and fibronectin |
| AAEL000726-RA | 3.6985 | 4.1 × 10−42 | fibrinogen and fibronectin |
| AAEL003894-RA | −1.8649 | 3.0 × 10−3 | Gram-Negative Binding Protein (GNBP) |
| AAEL003966-RA | −1.9430 | 2.8 × 10−6 | lachesin |
| AAEL010125-RA | 2.3599 | 8.2 × 10−10 | leucine-rich immune protein (Coil-less) |
| AAEL010132-RA | 2.4747 | 2.5 × 10−5 | leucine-rich immune protein (Long) |
| AAEL012255-RA | 1.8045 | 7.8 × 10−4 | leucine-rich immune protein (Short) |
| AAEL001420-RA | 1.9602 | 5.0 × 10−10 | leucine-rich immune protein (Short) |
| AAEL001401-RA | 2.0463 | 3.8 × 10−18 | leucine-rich immune protein (Short) |
| AAEL010772-RA | −2.2935 | 3.6 × 10−6 | leucine-rich repeat-containing protein |
| AAEL011760-RA | −2.2672 | 9.7 × 10−4 | leucine-rich transmembrane protein |
| AAEL012093-RA | 2.0559 | 2.0 × 10−4 | leucine-rich transmembrane protein |
| AAEL008271-RA | 6.5329 | 4.5 × 10−12 | SEC14, putative protein |
| AAEL007432-RA | 6.4799 | 1.5 × 10−65 | serine collagenase 1 precursor |
| AAEL002510−RB | 1.8323 | 1.4 × 10−4 | serine hydro×ymethyltransferase |
| AAEL000224-RA | −3.2320 | 2.1 × 10−3 | serine protease |
| AAEL013427-RA | −3.1475 | 4.8 × 10−4 | serine protease |
| AAEL007106-RA | 2.1779 | 2.0 × 10−4 | serine protease |
| AAEL002704-RB | 3.1322 | 3.4 × 10−37 | Serine Protease Inhibitor (serpin) |
| AAEL002731-RA | 2.1521 | 2.7 × 10−10 | Serine Protease Inhibitor (serpin) |
| AAEL007420-RB | 3.3676 | 6.7 × 10−23 | Serine Protease Inhibitor (serpin) |
| AAEL003182-RA | 3.5677 | 2.1 × 10−43 | Serine Protease Inhibitor (serpin) |
| AAEL011891-RA | 2.7316 | 6.5 × 10−3 | serine-type endopeptidase |
| AAEL006902-RA | 3.1972 | 6.8 × 10−6 | serine-type endopeptidase |
| AAEL008567-RA | 3.5171 | 5.4 × 10−13 | serine-type endopeptidase |
| AAEL001703-RA | 3.7862 | 2.8 × 10−8 | serine-type endopeptidase |
| AAEL008784-RA | 3.9399 | 6.6 × 10−6 | serine-type endopeptidase |
| AAEL001674-RA | 5.4452 | 6.4 × 10−6 | serine-type endopeptidase |
| AAEL014188-RA | 5.5757 | 1.8 × 10−9 | serine-type endopeptidase |
| AAEL009843-RA | 5.7334 | 2.4 × 10−59 | serine-type endopeptidase |
| AAEL013284-RA | 5.8404 | 1.4 × 10−8 | serine-type endopeptidase |
| AAEL003060-RA | 6.2886 | 7.2 × 10−129 | serine-type endopeptidase |
| AAEL001693-RA | 7.8599 | 1.2 × 10−109 | serine-type endopeptidase |
| AAEL001690-RA | 8.1855 | 7.1 × 10−114 | serine-type endopeptidase |
| AAEL008236-RA | −2.6394 | 9.4 × 10−3 | sidestep protein |
| AAEL000398-RA | −2.4779 | 4.8 × 10−3 | sidestep protein |
| AAEL009943-RA | −2.2813 | 5.3 × 10−5 | sidestep protein |
| AAEL010645-RA | 3.8037 | 9.9 × 10−3 | sidestep protein |
| AAEL011989-RB | 1.9061 | 4.9 × 10−10 | signal peptide peptidase |
| AAEL014755-RA | −2.4382 | 1.7 × 10−3 | tep2 protein |
| AAEL008607-RA | −1.8312 | 4.5 × 10−10 | tep3 protein |
| AAEL015018-RA | 2.2916 | 6.7 × 10−4 | toll protein |
| AAEL004000-RA | −2.9297 | 5.8 × 10−4 | Toll-like receptor |
| AAEL009551-RA | −2.8670 | 1.4 × 10−3 | Toll-like receptor |
| AAEL012780-RA | −5.2801 | 3.1 × 10−3 | trypsin |
| AAEL006414-RA | −2.6981 | 1.7 × 10−7 | trypsin |
| AAEL006430-RA | −2.6562 | 2.2 × 10−3 | trypsin |
| AAEL006429-RA | −2.4442 | 3.8 × 10−5 | trypsin |
| AAEL005764-RA | −2.1564 | 7.9 × 10−16 | trypsin |
| AAEL010202-RA | 1.9991 | 2.3 × 10−3 | trypsin |
| AAEL008093-RA | 2.6642 | 1.1 × 10−7 | trypsin |
| AAEL008085-RA | 4.4810 | 5.5 × 10−27 | trypsin |
| AAEL013703-RA | 5.2463 | 1.2 × 10−72 | trypsin |
| AAEL006425-RA | 5.2958 | 2.4 × 10−19 | trypsin |
| AAEL013715-RA | 8.8303 | 1.9 × 10−42 | trypsin |
| AAEL007601-RA | 9.5930 | 1.2 × 10−90 | trypsin |
| AAEL013707-RA | 9.8853 | 1.9 × 10−50 | trypsin |
| AAEL013714-RA | 10.627 | 4.3 × 10−48 | trypsin |
| AAEL010196-RA | 10.743 | 1.5 × 10−36 | trypsin |
| AAEL007818-RB | 2.8577 | 9.9 × 10−7 | Trypsin 3A1 Precursor |
| AAEL013712-RA | 6.7673 | 1.2 × 10−24 | Trypsin 5G1 Precursor |
| AAEL013629-RA | 3.3504 | 2.1 × 10−13 | trypsin-alpha |
| AAEL008079-RA | 6.2776 | 1.7 × 10−16 | trypsin-alpha |
| AAEL006403-RA | −3.7628 | 4.8 × 10−10 | trypsin-beta |
| AAEL008080-RA | 2.3952 | 1.2 × 10−5 | trypsin-eta |
| AAEL013628-RA | 4.8704 | 4.2 × 10−14 | trypsin-eta |
| AAEL000793-RA | 3.3944 | 1.1 × 10−48 | venom allergen |
| Transcript ID | Log2 FC | p-adj | Gene Description |
|---|---|---|---|
| AAEL004223-RA | 1.8947 | 1.1 × 10−6 | cecropin anti-microbial peptide |
| AAEL000037-RA | 3.0672 | 1.4 × 10−3 | Clip-Domain Serine Protease family B |
| AAEL006161-RB | 3.1447 | 1.1 × 10−3 | Clip-Domain Serine Protease family B |
| AAEL003632-RA | 3.0778 | 6.0 × 10−3 | Clip-Domain Serine Protease family B |
| AAEL005641-RA | 3.2355 | 4.9 × 10−4 | C-Type Lectin (CTL)—galactose binding |
| AAEL011455-RA | 4.1030 | 4.2 × 10−3 | C-Type Lectin (CTL)—mannose binding |
| AAEL012353-RA | 4.3829 | 1.3 × 10−3 | C-Type Lectin (CTL) |
| AAEL003857-RA | 7.3899 | 4.1 × 10−6 | defensin anti-microbial peptide |
| AAEL006691-RA | 1.9004 | 8.2 × 10−3 | fibrinogen and fibronectin |
| AAEL001401-RA | 2.7153 | 3.9 × 10−4 | leucine-rich immune protein (Short) |
| AAEL005734-RA | −1.9928 | 2.9 × 10−14 | leucine-rich transmembrane protein |
| AAEL002295-RA | 2.2469 | 6.8 × 10−3 | leucine-rich transmembrane protein |
| AAEL003597-RB | 2.4398 | 1.6 × 10−3 | leucine-rich transmembrane protein |
| AAEL005844-RA | −2.2151 | 9.8 × 10−25 | posF21, putative protein |
| AAEL008271-RA | 3.0341 | 8.6 × 10−3 | SEC14 putative protein |
| AAEL003896-RA | −1.7801 | 2.4 × 10−27 | serine/threonine protein kinase |
| AAEL009244-RA | 1.8418 | 2.3 × 10−7 | serine-type endopeptidase |
| AAEL001703-RA | 2.0534 | 2.9 × 10−4 | serine-type endopeptidase |
| AAEL009843-RA | 2.8570 | 8.1 × 10−3 | serine-type endopeptidase |
| AAEL003060-RA | 2.3460 | 1.1 × 10−5 | serine-type endopeptidase |
| AAEL007818-RB | 2.5953 | 7.1 × 10−4 | Trypsin 3A1 Precursor |
| AAEL006425-RA | 2.3896 | 3.0 × 10−9 | trypsin |
| AAEL013703-RA | 2.5507 | 3.3 × 10−4 | trypsin |
| AAEL008080-RA | 2.7631 | 2.1 × 10−4 | trypsin-eta |
| AAEL012473-RB | −2.1156 | 9.4 × 10−6 | vav1 protein |
| Transcript ID | Log2 FC | p-adj | Gene Description |
|---|---|---|---|
| AAEL004223-RA | 2.3577 | 4.4 × 10−5 | cecropin anti-microbial peptide |
| AAEL017211-RA | 2.3359 | 3.3 × 10−4 | cecropin anti-microbial peptide |
| AAEL009680-RB | 4.7006 | 1.6 × 10−10 | chymotrypsin |
| AAEL000556-RA | 4.0712 | 6.3 × 10−6 | C-Type Lectin (CTL) |
| AAEL000533-RA | 3.0535 | 9.0 × 10−4 | C-Type Lectin (CTL) |
| AAEL009670-RA | 3.6210 | 1.3 × 10−3 | C-Type Lysozyme (Lys-D) |
| AAEL003849-RA | 3.1292 | 4.3 × 10−3 | defensin anti-microbial peptide |
| AAEL000726-RA | 4.7815 | 5.9 × 10−8 | fibrinogen and fibronectin |
| AAEL006691-RA | 3.2048 | 2.7 × 10−6 | fibrinogen and fibronectin |
| AAEL007942-RA | 3.4734 | 7.6 × 10−6 | fibrinogen and fibronectin |
| AAEL011007-RA | 4.5979 | 1.0 × 10−6 | fibrinogen and fibronectin |
| AAEL004522-RA | 3.1695 | 2.6 × 10−3 | gambicin anti-microbial peptide |
| AAEL012092-RA | 2.0567 | 8.4 × 10−3 | leucine-rich repeat |
| AAEL002295-RA | 2.6054 | 1.3 × 10−4 | leucine-rich transmembrane protein |
| AAEL007420-RB | 5.1106 | 2.4 × 10−5 | Serine Protease Inhibitor (serpin) |
| AAEL003182-RA | 4.6609 | 7.4 × 10−4 | Serine Protease Inhibitor (serpin) |
| AAEL002704-RB | 4.2747 | 1.5 × 10−3 | Serine Protease Inhibitor (serpin) |
| AAEL006902-RA | 3.6563 | 3.6 × 10−6 | serine-type endopeptidase |
| AAEL008567-RA | 3.1476 | 7.6 × 10−9 | serine-type endopeptidase |
| AAEL008784-RA | 3.6928 | 1.4 × 10−4 | serine-type endopeptidase |
| AAEL009244-RA | 3.0981 | 4.5 × 10−3 | serine-type endopeptidase |
| AAEL003060-RA | 2.2077 | 3.1 × 10−4 | serine-type endopeptidase |
| AAEL007818-RB | 4.8704 | 5.5 × 10−3 | Trypsin 3A1 Precursor |
| AAEL007818-RA | 4.1455 | 3.2 × 10−5 | Trypsin 3A1 Precursor |
| AAEL006425-RA | 3.2268 | 1.5 × 10−10 | trypsin |
| AAEL008085-RA | 2.7895 | 2.4 × 10−4 | trypsin |
| AAEL013623-RA | 3.0234 | 9.7 × 10−4 | trypsin |
| AAEL013703-RA | 2.7960 | 7.2 × 10−13 | trypsin |
| AAEL008079-RA | 2.9739 | 1.9 × 10−10 | trypsin-alpha |
| AAEL013628-RA | 4.2736 | 7.2 × 10−13 | trypsin-eta |
| AAEL000793-RA | 4.7662 | 6.8 × 10−5 | venom allergen |
| AAEL002693-RA | 2.4160 | 9.6 × 10−3 | venom allergen |
| Transcript ID | Log2 FC | p-adj | Gene Description |
|---|---|---|---|
| AAEL010235-RA | −5.6356 | 4.0 × 10−18 | allergen |
| AAEL006424-RA | −3.1799 | 3.1 × 10−4 | allergen |
| AAEL001139-RA | −3.7087 | 9.1 × 10−3 | br serine/threonine-protein kinase |
| AAEL017211-RA | −2.0884 | 1.4 × 10−4 | cecropin anti-microbial peptide |
| AAEL005374-RA | −3.5005 | 2.6 × 10−4 | Class B Scavenger Receptor (CD36 domain) |
| AAEL000234-RA | −2.1910 | 2.4 × 10−3 | Class B Scavenger Receptor (CD36 domain) |
| AAEL006355-RA | 2.9459 | 5.5 × 10−5 | Class B Scavenger Receptor |
| AAEL003628-RA | 2.5283 | 2.5 × 10−4 | Clip-Domain Serine Protease family B |
| AAEL011612-RB | −3.1179 | 1.0 × 10−2 | C-Type Lectin (CTL)—mannose binding |
| AAEL000556-RA | −2.9238 | 1.4 × 10−6 | C-Type Lectin (CTL) |
| AAEL006417-RA | −4.6920 | 2.9 × 10−9 | D7 protein |
| AAEL000726-RA | −3.3941 | 9.3 × 10−8 | fibrinogen and fibronectin |
| AAEL007942-RA | −2.4764 | 1.1 × 10−4 | fibrinogen and fibronectin |
| AAEL011007-RA | −2.9856 | 1.3 × 10−4 | fibrinogen and fibronectin |
| AAEL007064-RA | −2.7633 | 2.4 × 10−3 | Gram-Negative Binding Protein (GNBP) |
| AAEL000576-RA | 3.8061 | 1.3 × 10−3 | lachesin |
| AAEL012092-RA | −2.0902 | 1.7 × 10−3 | leucine-rich repeat protein |
| AAEL000243-RA | 4.4869 | 7.5 × 10−4 | leucine-rich transmembrane protein |
| AAEL003597-RB | 2.3056 | 2.3 × 10−3 | leucine-rich transmembrane protein |
| AAEL007420-RB | −3.1285 | 1.3 × 10−4 | Serine Protease Inhibitor (serpin) |
| AAEL003182-RA | −3.8025 | 4.7 × 10−4 | Serine Protease Inhibitor (serpin) |
| AAEL002704-RB | −4.2338 | 4.3 × 10−5 | Serine Protease Inhibitor (serpin) |
| AAEL010267-RA | 4.6569 | 5.4 × 10−3 | serine protease |
| AAEL003508-RB | −2.5065 | 1.0 × 10−9 | serine-pyruvate aminotransferase |
| AAEL001701-RA | −3.9779 | 6.3 × 10−3 | serine-type endopeptidase |
| AAEL008271-RA | 6.5329 | 8.8 × 10−3 | toll pathway signaling |
| AAEL007432-RA | 6.4799 | 6.8 × 10−3 | toll protein |
| AAEL002510-RB | 1.8323 | 1.2 × 10−3 | Toll-like receptor |
| AAEL000224-RA | −3.2320 | 5.4 × 10−3 | Toll-like receptor |
| AAEL007992-RA | −2.0267 | 2.4 × 10−3 | trypsin |
| AAEL005596-RA | −3.6009 | 1.1 × 10−5 | trypsin-epsilon |
| AAEL000793-RA | −3.7994 | 5.4 × 10−5 | venom allergen |
| AAEL006297-RA | −2.6718 | 1.3 × 10−3 | venom allergen |
| Transcript ID | Log2 FC | p-adj | Gene Description |
|---|---|---|---|
| AAEL013063-RA | 1.9928 | 9.8 × 10−8 | autophagy related gene |
| AAEL009384-RA | −3.7894 | 4.5 × 10−3 | fibrinogen and fibronectin |
| AAEL009178-RA | −3.8119 | 2.1 × 10−4 | Gram-Negative Binding Protein (GNBP) |
| AAEL009692-RA | 1.8392 | 2.1 × 10−3 | JAKSTAT pathway signaling signal |
| AAEL005687-RA | −2.6650 | 2.9 × 10−3 | protein serine/threonine kinase |
| AAEL001914-RA | −2.0191 | 2.7 × 10−3 | scavenger receptor |
| AAEL010267-RA | 5.0054 | 8.3 × 10−6 | serine protease |
| AAEL007835-RA | −2.2677 | 1.1 × 10−5 | serine/threonine protein kinase |
| AAEL014188-RA | −2.6299 | 2.4 × 10−5 | serine-type endopeptidase |
| AAEL001701-RA | −3.7945 | 7.5 × 10−3 | serine-type endopeptidase |
| AAEL007992-RA | −3.8714 | 3.2 × 10−3 | trypsin |
| AAEL008080-RA | −2.7126 | 4.9 × 10−3 | trypsin-eta |
| AAEL010486-RA | −2.2017 | 3.9 × 10−4 | viral IAP-associated factor |
| Treatment | Pillai’s Trace | Degrees of Freedom (Numerator, Denominator) | p-Value | Standardized Canonical Coefficients | |||||
|---|---|---|---|---|---|---|---|---|---|
| AaeDEFA | AaeDEFD | AaeDEFa | AaeDNR1 | AaeCECH | AaeTEP3 | ||||
| Strain | 0.95 | 6, 23 | <0.0001 | 2.26 | 0.53 | −1.79 | −0.29 | 0.45 | 4.48 |
| Time | 4.05 | 36, 168 | <0.0001 | 4.45 | 2.38 | −0.77 | −0.39 | 1.39 | −2.34 |
| Strain × Time | 3.88 | 36, 168 | <0.0001 | 5.37 | 2.95 | −0.97 | −0.12 | −1.57 | 2.47 |
| Gene ID | Accession | Gene Name | Primer Name | Primer Sequence (5’–3’) |
|---|---|---|---|---|
| AAEL011197 | XM_001655125 | Actin | AaeActin-197-152F | AGGACTCGTACGTCGGTGAC |
| AaeActin-197-590R | CGTTCAGTCAGGATCTTC | |||
| AAEL003849 1 | XM_021856546 | DEFA | AaeDEF-A-849-208F | CGCCCTTTTGCAAACTCTCT |
| AaeDEF-A-849-380R | TTGCAGTAACCTCCCCGATT | |||
| AAEL003857 | XM_001657239 | DEFD | AaeDEF-D-857-39F | CACCGGGGCCATTACTAGTG |
| AaeDEF-D-857-196R | CGCTCAACAGATCACAGGTG | |||
| AAEL003841 | XM_001657243 | DEFa | AaeDEF-A-841-208F | CGCCCTTTTGCAAACTCTCT |
| AaeDEF-A-841-380R | TTGCAGTAACCTCCCCGATT | |||
| AAEL017211 | NW_001809913 | CECH | AaeCEC-211-131F | CAAGCTGCTATTGGTGGTCG |
| AaeCEC-211-312R | CGTTCACGCTTGTCTAAACCA | |||
| AAEL008607 | XM_021846500 | tep3 | AaeTep3-607-684F | AGTGTCCGTTGAGTCTCCTG |
| AaeTep3-607-890R | TCTACCGATCCCTTGCCATC | |||
| AAEL000590 | XM_001648620 | DNR1 | AaeDNR1-590-550F | AGCATTGCATCGACAGTCAC |
| AaeDNR1-590-738R | AGCGGAACTTGCAGTCATTT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Alto, B.W.; Jiang, Y.; Yu, F.; Zhang, Y. Transcriptomic Analysis of Aedes aegypti Innate Immune System in Response to Ingestion of Chikungunya Virus. Int. J. Mol. Sci. 2019, 20, 3133. https://doi.org/10.3390/ijms20133133
Zhao L, Alto BW, Jiang Y, Yu F, Zhang Y. Transcriptomic Analysis of Aedes aegypti Innate Immune System in Response to Ingestion of Chikungunya Virus. International Journal of Molecular Sciences. 2019; 20(13):3133. https://doi.org/10.3390/ijms20133133
Chicago/Turabian StyleZhao, Liming, Barry W. Alto, Yongxing Jiang, Fahong Yu, and Yanping Zhang. 2019. "Transcriptomic Analysis of Aedes aegypti Innate Immune System in Response to Ingestion of Chikungunya Virus" International Journal of Molecular Sciences 20, no. 13: 3133. https://doi.org/10.3390/ijms20133133
APA StyleZhao, L., Alto, B. W., Jiang, Y., Yu, F., & Zhang, Y. (2019). Transcriptomic Analysis of Aedes aegypti Innate Immune System in Response to Ingestion of Chikungunya Virus. International Journal of Molecular Sciences, 20(13), 3133. https://doi.org/10.3390/ijms20133133